
Comparación de cráneos de distintas especies tempranas de Homo. Chris Stringer, Natural History Museum, United Kingdom – Stringer, Chris (años 10eptember 2015). «The many mysteries of Homo naledi«. CC BY 4.0.
Se comparan réplicas de cráneos de (de izquierda a derecha) Homo habilis (KNM-ER 1813, Koobi Fora, Kenia, aproximadamente 1.8 millones de años), un Homo erectus temprano (D2700, Dmanisi, Georgia, aproximadamente 1.8 millones de años) y Homo floresiensis (Liang Bua 1, Indonesia, aproximadamente 20,000 años) con fragmentos reales de material craneal de Homo naledi que se han superpuesto en una reconstrucción virtual (extremo derecho; observe que algunas de las imágenes del material de H. naledi han sido invertidas). En cada caso, los cráneos están etiquetados con las características típicas de cada especie. Por ejemplo, mientras que el volumen cerebral de los humanos modernos (Homo sapiens) adultos es típicamente entre 1000 y 1500 centímetros cúbicos (cc), H. habilis variaba de aproximadamente 510 a >700 cc, H. erectus de aproximadamente 550 a >1100 cc, H. floresiensis alrededor de 426 cc, y H. naledi entre 466 y 560 cc. Además, en los humanos modernos, el hueso occipital (en la parte posterior del cráneo) típicamente tiene una forma redondeada en el perfil, mientras que en algunos humanos primitivos como H. erectus, las porciones superior e inferior del occipital están anguladas entre sí (es decir, «flexionadas»), y hay una cresta ósea pronunciada que atraviesa la región angulada (llamada toro transversal).
Los homininos son el grupo de primates al que pertenecemos los seres humanos. En ellos se inicia una transformación decisiva dentro de la evolución: el paso de una vida arbórea a una existencia cada vez más terrestre, marcada por la postura erguida y la marcha bípeda. No se trata solo de un cambio físico, sino de una nueva forma de relación con el entorno.
Desde el punto de vista científico, los homininos constituyen una subtribu (Hominina) dentro de la familia de los homínidos. Hoy solo sobrevive una especie, Homo sapiens, pero durante millones de años existieron otras formas humanas que compartieron rasgos comunes y que fueron dando forma, lentamente, a nuestra propia especie.
Si se aceptan especies como Orrorin tugenensis o Sahelanthropus tchadensis, el origen de los homininos podría situarse hace unos seis o siete millones de años. En ese momento debió de existir el antepasado común del ser humano y el chimpancé, a partir del cual se separaron sus respectivas líneas evolutivas.
Los homininos se caracterizan por una adaptación progresiva a la vida en el suelo. La postura erguida implica una reorganización completa del cuerpo: el cráneo se equilibra sobre la columna, la pelvis cambia de forma y los pies dejan de ser prensiles. El dedo gordo se alinea con los demás, permitiendo una marcha más estable. Al mismo tiempo, las manos, con un pulgar desarrollado, se especializan en la manipulación de objetos, abriendo el camino hacia el uso de herramientas.
Historia paleontológica
El estudio de los homininos se apoya en un conjunto de hallazgos fósiles localizados, en su mayor parte, en el este de África. Allí, a lo largo del Gran Valle del Rift —una enorme fractura de la corteza terrestre que se extiende desde Mozambique hasta el mar Muerto—, se han conservado algunos de los restos más antiguos de nuestra historia evolutiva.
La formación de este gran sistema geológico estuvo acompañada de una intensa actividad volcánica. Las capas de cenizas y materiales piroclásticos generadas en ese contexto han resultado fundamentales para la ciencia, ya que permiten datar los fósiles con bastante precisión mediante métodos como el potasio-argón o el paleomagnetismo. Gracias a ello, es posible situar los restos dentro de una cronología relativamente fiable.
Desde el punto de vista biológico, los chimpancés son nuestros parientes más cercanos. Compartimos con ellos aproximadamente el 98 % del material genético. A partir de estos datos, la biología molecular ha estimado que la línea evolutiva de los homininos se separó de la de los chimpancés hace entre seis y siete millones de años.
Este cálculo se basa en los llamados «relojes moleculares», que relacionan el grado de diferencia genética entre dos especies con el tiempo transcurrido desde su separación. Aunque se trata de una estimación, resulta coherente con los cambios ambientales que tuvieron lugar en África en ese periodo.
Durante el final del Mioceno, importantes transformaciones climáticas provocaron la reducción de las grandes selvas tropicales y la expansión de paisajes más abiertos, dominados por sabanas. Este nuevo entorno favoreció la desaparición de algunas especies y la aparición de otras adaptadas a condiciones diferentes, entre ellas los primeros homininos.
Durante años, uno de los homininos más antiguos conocidos fue Ardipithecus ramidus, descubierto entre 1994 y 1997 por el equipo de Tim White en la región etíope de Afar. Esta especie, con una antigüedad aproximada de 4,4 millones de años, habitaba entornos boscosos y presentaba una alimentación similar a la de los chimpancés. Su tamaño era reducido, en torno a un metro de altura y unos treinta kilogramos de peso.
Algunos rasgos anatómicos, como la reducción de los caninos o la posición de la base del cráneo, sugieren una forma primitiva de bipedismo. Este planteamiento se vio reforzado por el hallazgo posterior de restos óseos compatibles con la locomoción bípeda. No obstante, su forma de desplazarse debió de ser distinta a la de cualquier especie actual.
En un primer momento, se llegó a considerar a Ardipithecus ramidus como un posible «eslabón perdido». Sin embargo, hoy se tiende a interpretarlo como una rama lateral dentro del complejo árbol evolutivo humano.
A comienzos del siglo XXI se identificaron restos aún más antiguos en Etiopía, datados entre 5,2 y 5,8 millones de años, que fueron atribuidos a Ardipithecus kadabba. Esta especie presenta algunas similitudes dentales con Orrorin tugenensis, lo que sugiere la existencia de una diversidad mayor de formas tempranas de homininos de lo que se pensaba inicialmente.
Ver: (www.portalciencia.net Ardipithecus kadabba 3 de junio de 2008).
Reconstrucción de Australopithecus. Foto: Wolfgang Sauber – Trabajo propio. Natural History Museum, Vienna ( Austria ). Model of a male Australopithecus afarensis.CC BY-SA 4.0.

En diciembre de 2000, Brigitte Senut y Martin Pickford anunciaron el hallazgo de unos fósiles muy antiguos en las colinas de Tugen, cerca del lago Baringo, en Kenia. A este descubrimiento se le dio el nombre popular de «ancestro del milenio», aunque su denominación científica es Orrorin tugenensis. Algunos restos del fémur sugieren que este ser habría practicado la locomoción bípeda. Su antigüedad, situada entre 5,9 y 6,1 millones de años, lo convierte en uno de los candidatos más antiguos dentro de la línea de los homininos. Este hallazgo podría guardar relación con un fragmento de mandíbula descubierto en 1967 en Lothagam, también en Kenia, que en su momento dio lugar a una notable controversia.
En 2002, el equipo de Michel Brunet presentó un cráneo de unos 350 cm³ de capacidad, con una antigüedad de entre 6 y 7 millones de años. El fósil, hallado en el norte del Chad, mostraba rasgos en parte similares a los del chimpancé, aunque su cara presentaba características más próximas a las humanas. El ejemplar recibió el nombre popular de Toumaï, mientras que la especie fue denominada Sahelanthropus tchadensis. Aunque también se han encontrado restos de las extremidades, la posición del foramen magno en la base del cráneo ha sido uno de los argumentos utilizados para defender que podía tratarse de un ser bípedo.
La inclusión de Sahelanthropus tchadensis entre los homininos sigue siendo objeto de debate. De hecho, Senut y Pickford no aceptan esta interpretación, mientras que los descubridores de Ardipithecus han sugerido que podría tratarse de una forma muy arcaica próxima a ese género.
En abril de 2005, Brunet y sus colaboradores presentaron una reconstrucción virtual del cráneo que reforzaba la hipótesis de su inclusión entre los homininos. Aun así, la falta de restos poscraneales más completos impide alcanzar una certeza definitiva.
Evolución
Actualmente se considera que, hace unos tres millones de años, a partir de los australopitecos se abrieron dos grandes líneas evolutivas paralelas. Por un lado apareció el género Paranthropus, formado por especies más robustas y especializadas en una dieta vegetal dura, compuesta por tubérculos, tallos y nueces. Esa alimentación exigía una mandíbula poderosa, molares de gran tamaño y una musculatura facial muy desarrollada, en ocasiones asociada a una cresta sagital en el cráneo.
Por otro lado surgió la línea del género Homo, caracterizada por un cerebro relativamente mayor, una mandíbula y unos dientes más reducidos, y una constitución ósea más grácil. Durante alrededor de un millón de años, ambos géneros coexistieron en África, representando dos formas distintas de adaptación dentro de la evolución de los homininos.
Casi al mismo tiempo que Paranthropus aethiopicus apareció otro hominino con un encéfalo más desarrollado: Homo habilis. A esta especie se le atribuye de manera tradicional la cultura de los guijarros tallados, una de las manifestaciones más antiguas de tecnología lítica. En un primer momento recibió nombres como Telanthropus o Prezinjantropus, reflejo de las dudas iniciales sobre su clasificación.
Homo habilis suele interpretarse como una forma intermedia entre los australopitecos y Homo ergaster, aunque algunos autores prefieren distinguir dentro de este grupo a Homo rudolfensis como especie separada. Su capacidad craneal, situada entre 600 y 700 cm³, era claramente superior a la de los australopitecos. Sus restos se han hallado en diversos puntos del África oriental, lo que indica una expansión considerable dentro de esa región.
Cráneo de Tautavel. Tautavel Man (Homo erectus tautavelensis) from 450 kya Arago Cave, France. CC BY 2.5.
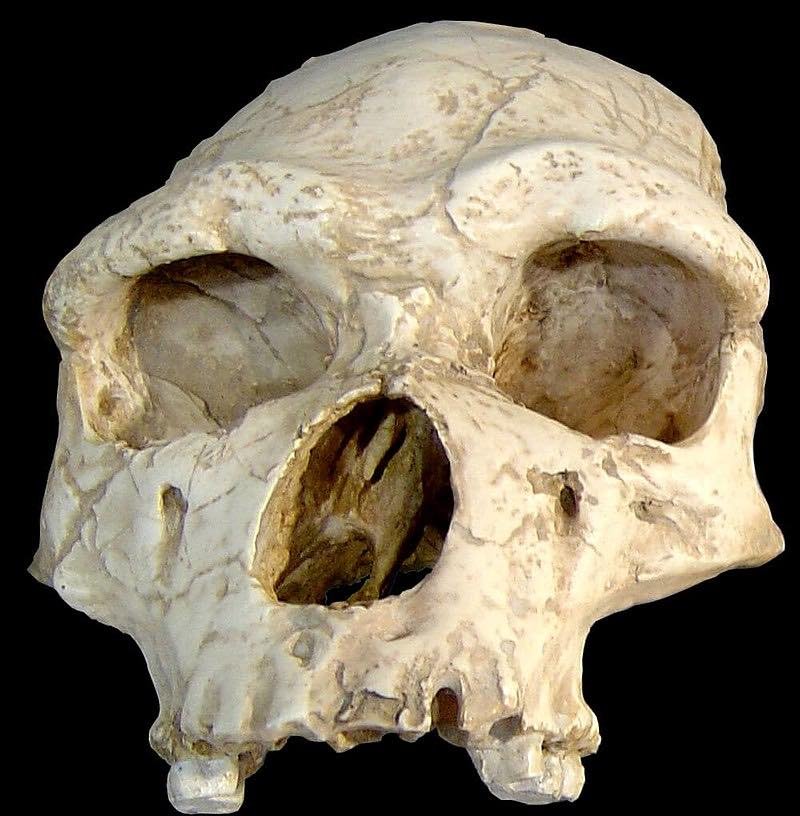
La expresión hombre de Tautavel designa un conjunto de fósiles de homínidos que datan de cerca de 450.000 años AP, encontrados en la cueva de l’Arago, Tautavel, Pirineos Orientales, por el equipo de Enrique de Lumley.
La industria lítica asociada al Hombre de Tautavel se interpreta unas veces como un Tayaciense antiguo, y otras como un Achelense. Hay algunos bifaces, pero son raros habida cuenta la dificultad de ser realizados sobre los materiales locales.
Parece que el hombre de Tautavel no dominaba aún el fuego: pocas osamentas quemadas que certifican la utilización del fuego en Caune de l’Arago, aparecen en depósitos posteriores, cuya antigüedad está entre los 400.000 y 100.000 años AP, y los testimonios de fogatas solo se generalizan a partir de 100.000 años AP.
El hábitat reveló restos de rinocerontes y caballos. Es muy posible que comiera también animales pequeños. Algunos investigadores formularon la hipótesis de que era caníbal y más probablemente carroñero, más que cazador. Si fue cazador, su territorio se extendió seguramente sobre un radio de 30 km (como prueban también algunas rocas importadas para fabricar sus herramientas).
El Primer Homínido que controló el fuego | Homo Erectus
Homo erectus es un homínido extinto que vivió entre 1,9 millones de años y 117 000 años antes del presente. El cuerpo era alto, podía medir hasta 1,80 m, muy robusto, esbelto y moderno, morfológicamente similar al humano actual. Producía industria lítica, principalmente Achelense, y probablemente dominaba el fuego…
Evolución posterior del género Homo
Homo ergaster, considerado probablemente descendiente de Homo habilis, fue una de las primeras especies humanas con gran éxito adaptativo. Surgió en África y, al parecer, protagonizó una de las primeras expansiones fuera de este continente hace al menos 1,8 millones de años. Restos atribuidos a formas emparentadas con este proceso se han hallado en Georgia, China, Java y otros lugares de Asia, lo que muestra una temprana difusión de los homininos por amplias regiones del Viejo Mundo.
En ese contexto, algunos autores distinguen formas como Homo georgicus, mientras que en Asia oriental acabó consolidándose Homo erectus, una de las especies más duraderas y exitosas de la evolución humana. Mucho más tarde, en el ámbito insular del sudeste asiático, pudo surgir Homo floresiensis, una pequeña especie humana adaptada a condiciones muy particulares.
En el extremo occidental de Eurasia, la evolución de estas poblaciones dio lugar a formas humanas antiguas cuyos restos se han encontrado en Atapuerca, en España. Entre ellas destaca Homo antecessor, documentado en niveles de gran antigüedad, en torno a 1,2 millones de años. Su posición exacta dentro del árbol evolutivo sigue siendo debatida, aunque a menudo se le ha relacionado con fases tempranas de la presencia humana en Europa.
Más adelante apareció Homo heidelbergensis, especie a la que muchos investigadores consideran próxima al origen de los neandertales en Europa. A su vez, los neandertales desarrollaron una trayectoria propia y llegaron a ocupar gran parte del continente europeo y zonas de Asia occidental durante el Pleistoceno. Poseían una capacidad craneal comparable, e incluso en ocasiones superior, a la del ser humano actual.
Mientras tanto, en África evolucionaron otras poblaciones humanas descendientes de formas próximas a Homo ergaster o emparentadas con ellas. Dentro de este marco se ha utilizado el nombre Homo rhodesiensis para designar algunos fósiles africanos de hace entre 500 000 y 300 000 años, que podrían situarse cerca del origen de Homo sapiens. No obstante, la clasificación exacta de estos restos sigue siendo objeto de discusión.
Homo sapiens surgió en África, probablemente en regiones orientales y meridionales del continente, y desde allí se expandió primero por África y después por Eurasia, Oceanía y, mucho más tarde, por América. La genética ha permitido reconstruir de forma cada vez más precisa estas migraciones y ha mostrado que la humanidad actual procede de poblaciones africanas relativamente recientes en términos evolutivos.
También sabemos hoy que los neandertales no fueron antepasados directos de los sapiens, sino una humanidad paralela, con su propia historia evolutiva. Sin embargo, ambas especies llegaron a encontrarse y a mezclarse en algunos lugares, de modo que una pequeña parte de la herencia genética neandertal sigue presente en muchas poblaciones humanas actuales.
Referencias
Notas
- www.portalciencia.net Ardipithecus kadabba 3 de junio de 2008
- Analyzing Hominin Phylogeny: Cladistic Approach. Link Springer.
Bibliografía
- Mann, Alan and Mark Weiss 1996: «Hominoid Phylogeny and Taxonomy: a consideration of the molecular and Fossil Evidence in an Historical Perspective.» Molecular Phylogenetics and Evolution 5(1): 169-181.
- SPOTORNO AE. 2014b. Capítulo 15 «Genética y Evolución». Pp. 215 .. en S. Berríos (ed.), «Genética Humana¨. 2014. Ed. Mediterráneo, Santiago de Chile
Género Homo
Homo (del latín homo, «hombre» o «ser humano») es el género al que pertenece nuestra especie, Homo sapiens. Se integra dentro de los primates homínidos, en la tribu Hominini, y reúne a las especies humanas actuales y extinguidas más próximas entre sí dentro de la evolución reciente de los homininos.
Los miembros del género Homo se caracterizan por una locomoción bípeda plenamente desarrollada, con pies no prensiles y el dedo gordo alineado con los demás, así como por una postura corporal completamente erguida. A ello se añade un mayor desarrollo del encéfalo y una reorganización del cráneo, rasgos que distinguen a este grupo de formas anteriores.
En líneas generales, el origen del género Homo se sitúa hace unos 2,5 millones de años, normalmente en relación con especies como Homo habilis y, según algunos autores, también Homo rudolfensis. A lo largo del tiempo, este género incluyó diversas especies humanas que se extendieron por África, Eurasia y algunas regiones insulares del sudeste asiático.
En la actualidad, la única especie viva es Homo sapiens. Las demás han desaparecido. Entre las supervivencias más tardías se suelen mencionar a Homo neanderthalensis en Europa, extinguido hace unos 40 000 años aproximadamente, y a Homo floresiensis en Indonesia, que habría perdurado hasta fechas relativamente recientes dentro de la prehistoria humana.
Uno de los criterios que llevó a separar a Homo habilis del género Australopithecus fue el aumento del tamaño cerebral, pero también la capacidad técnica. La fabricación de herramientas de piedra y, sobre todo, la posibilidad de conservarlas para utilizarlas más adelante, marcaron un cambio importante en la conducta y en la relación de estos homininos con su entorno.

Australopithecus garhi
Australopithecus garhi fue una especie de hominino que vivió en África oriental hace aproximadamente 2,5 millones de años, en un momento especialmente importante de la evolución humana. Sus restos fueron hallados en la región de Bouri, en el área de Middle Awash, en Etiopía, y su descripción científica se publicó a finales de la década de 1990. Su antigüedad lo sitúa en una fase de transición muy sugerente, cerca del periodo en el que comenzaron a aparecer las primeras formas del género Homo. Precisamente por eso ha despertado tanto interés entre los paleoantropólogos: porque pertenece a una época en la que el árbol evolutivo humano estaba diversificándose y en la que varias líneas de homininos coexistían en África.
El nombre garhi procede de una palabra de la lengua afar que significa algo parecido a “sorpresa”, y no es un nombre casual. Cuando se dieron a conocer sus restos, esta especie llamó la atención porque combinaba rasgos que recordaban a australopitecos más antiguos con otros que parecían anunciar cambios evolutivos de gran importancia. En otras palabras, no era una forma claramente moderna ni tampoco un simple representante más de los australopitecos clásicos. Presentaba una mezcla anatómica que lo convertía en una pieza especialmente interesante para estudiar cómo se pasó de los homininos tempranos a formas humanas posteriores.
Desde el punto de vista cronológico, Australopithecus garhi vivió en torno a esos 2,5 millones de años, una fecha muy significativa porque coincide con un periodo en el que África oriental estaba experimentando cambios ecológicos profundos. Los paisajes se estaban haciendo más abiertos y variables, alternando zonas de bosque, matorral y espacios de sabana. Esa transformación del medio debió de influir mucho en la evolución de los homininos, obligándolos a adaptarse a entornos más cambiantes, a desplazarse más y quizá también a diversificar su alimentación. A. garhi aparece precisamente en ese marco de transición ecológica, cuando la evolución humana parece acelerarse y multiplicar sus experimentos biológicos.
En cuanto a su aspecto, Australopithecus garhi conservaba varios rasgos propios de los australopitecos. Su cerebro seguía siendo pequeño en comparación con el de los humanos posteriores, con una capacidad craneal estimada en torno a 450 cm³, es decir, muy lejos todavía de las cifras del género Homo. Su cara era prognata, es decir, proyectada hacia delante, y sus dientes posteriores eran grandes y robustos, con esmalte grueso, algo frecuente entre los australopitecos. Sin embargo, algunos detalles de su anatomía han llevado a pensar que no se trataba de una forma cualquiera, sino de un hominino con características particulares dentro de este grupo.
Uno de los aspectos más comentados de A. garhi tiene que ver con su locomoción y con la proporción de sus miembros. Como otros homininos de su tiempo, era bípedo, pero algunos restos postcraneales sugieren una combinación anatómica compleja. Se ha señalado que el fémur muestra rasgos compatibles con la marcha bípeda, mientras que otros elementos del esqueleto recuerdan todavía una anatomía no del todo separada del mundo arborícola. Además, algunos investigadores han llamado la atención sobre la relativa longitud del fémur en comparación con otras partes del cuerpo, lo que ha sido interpretado como un posible indicio de cambios en el modo de desplazarse. No obstante, como ocurre a menudo en paleoantropología, estas interpretaciones deben manejarse con prudencia, porque el registro fósil es incompleto y no permite reconstrucciones absolutamente seguras.
La gran importancia de Australopithecus garhi no reside solo en su anatomía, sino en su posición temporal. Vivió justo en una franja del tiempo que durante años ha sido considerada crucial para entender el origen del género Homo. Por eso, cuando fue descrito, algunos investigadores propusieron que podía ser un candidato razonable a antepasado de las primeras especies humanas. La idea resultaba atractiva: un australopiteco relativamente tardío, con una mezcla peculiar de rasgos, situado casi en el umbral cronológico del género Homo. Sin embargo, con el paso del tiempo, la visión se ha vuelto más cauta. Hoy suele hablarse de A. garhi como una especie muy interesante y cercana a ese momento evolutivo decisivo, pero no como un antepasado directo demostrado de nuestra línea.
Otro motivo por el que esta especie ha sido muy discutida es su posible relación con las primeras herramientas de piedra y con el aprovechamiento de animales mediante corte. En la zona de Bouri aparecieron restos faunísticos con marcas que algunos investigadores interpretaron como señales de despiece con instrumentos líticos. Esto abrió la posibilidad de que homininos de esta época, tal vez Australopithecus garhi u otros muy próximos, ya estuvieran manipulando herramientas de piedra o utilizando objetos cortantes para aprovechar la carne y la médula de animales. Si esa relación fuese correcta, estaríamos ante una cuestión de enorme interés, porque la conducta técnica compleja no quedaría reservada exclusivamente al género Homo. Aun así, la prudencia también es necesaria aquí: la asociación entre fósiles, herramientas y marcas de corte no siempre permite afirmar con seguridad absoluta quién fue el autor de esas acciones.
En este sentido, Australopithecus garhi ocupa una posición muy sugerente dentro del panorama general de la evolución humana. No parece representar todavía a los humanos en sentido estricto, pero tampoco es una forma banal o marginal. Se sitúa en un momento en que África oriental albergaba varias líneas evolutivas de homininos, y en el que algunos rasgos anatómicos y conductuales estaban cambiando con rapidez. Su estudio ayuda a comprender que la evolución humana no fue una línea recta ni un relevo simple entre una especie y otra, sino un proceso de diversificación, coexistencia y ensayo evolutivo. Distintas poblaciones experimentaron soluciones diferentes ante los desafíos del medio, y solo algunas de esas líneas tendrían continuidad a largo plazo.
Por eso, cuando se habla de la antigüedad de Australopithecus garhi, no se trata solo de ponerle una cifra. Decir que vivió hace unos 2,5 millones de años significa situarlo en uno de los grandes umbrales de la prehistoria humana, cuando los australopitecos tardíos convivían con formas cada vez más próximas a los primeros humanos y cuando comenzaban a aparecer conductas técnicas que cambiarían para siempre la relación entre los homininos y su entorno. A. garhi pertenece todavía al mundo de los australopitecos, pero aparece ya muy cerca de la frontera donde empieza otra etapa.
En conjunto, Australopithecus garhi puede definirse como una especie africana tardía de australopiteco, conocida por restos hallados en Etiopía y datada en torno a 2,5 millones de años. Su combinación de rasgos primitivos y relativamente derivados, su cercanía cronológica al origen del género Homo y su posible asociación con actividades de corte o manipulación de herramientas la convierten en una figura clave para entender una fase especialmente compleja de la evolución humana. No es una especie famosa para el gran público como Lucy o los neandertales, pero dentro de la paleoantropología tiene un valor enorme, porque ilumina uno de los momentos más oscuros y apasionantes de nuestra historia remota.
Fuentes
Smithsonian Human Origins Program.
Asfaw et al., descripción científica de la especie en PubMed/Science.
Springer Nature Encyclopedia of Global Archaeology.
Artículo principal: Evolución humana
Evolución biológica del género Homo
La evolución biológica del género Homo constituye uno de los problemas más complejos y fascinantes de la paleoantropología. Durante mucho tiempo se intentó representar el origen humano como una secuencia lineal, ordenada y casi ascendente, en la que una especie daba paso con claridad a la siguiente. Sin embargo, el estudio de los fósiles hallados en África y Eurasia ha ido mostrando una realidad mucho más irregular. El nacimiento del aspecto humano no fue el resultado de una transformación única y uniforme, sino de un proceso largo, discontinuo y lleno de ramificaciones.
Uno de los mayores problemas para reconstruir este proceso es la existencia de importantes lagunas en el registro fósil, especialmente en el periodo comprendido entre hace unos dos y tres millones de años. Se trata de una etapa decisiva, porque en ella aparecen las primeras formas atribuidas al género Homo, pero también persisten otros homininos con rasgos más arcaicos. Esta convivencia de formas diferentes dificulta enormemente la reconstrucción de una línea evolutiva simple y obliga a pensar en un panorama mucho más abierto.
Hoy se acepta de forma bastante general que la evolución humana no siguió un camino recto. Más bien avanzó mediante una combinación de cambios anatómicos, conductuales y ecológicos que no siempre aparecieron al mismo tiempo ni en las mismas poblaciones. Por eso se habla a menudo de una evolución en mosaico: algunos rasgos considerados modernos surgieron antes que otros, y distintas especies compartieron combinaciones parciales de caracteres antiguos y recientes. En unas, el aumento del cerebro fue más visible; en otras, los cambios en la dentición, en la mano o en la locomoción resultaron más llamativos. El cuerpo humano, tal como hoy lo conocemos, no apareció de golpe, sino por ensamblaje gradual de transformaciones diversas.
La propia aparición del género Homo sigue siendo objeto de debate. Los fósiles más antiguos atribuidos a este género se sitúan en torno a 2,8–2,75 millones de años, pero el registro de esos primeros momentos es fragmentario y no permite trazar con seguridad una genealogía única. Algunas propuestas otorgan un papel importante a especies como Homo habilis o Homo rudolfensis, mientras que otras subrayan la dificultad de separar con nitidez a los primeros representantes de Homo de ciertos australopitecos tardíos. En este sentido, más que una frontera tajante entre un género y otro, parece haber existido una zona de transición en la que coexistieron rasgos mixtos y formas todavía inciertas.
A ello se suma otro factor importante: varias especies del género Homo pudieron convivir en una misma época. Esto rompe definitivamente la vieja imagen de una sucesión ordenada en la que una especie reemplazaba sin más a la anterior. En realidad, durante ciertos periodos coexistieron linajes distintos, con adaptaciones propias y trayectorias diferentes. Algunas poblaciones se expandieron fuera de África, otras permanecieron durante más tiempo en ecosistemas concretos, y otras desaparecieron sin dejar descendencia directa. La evolución humana se parece así menos a una escalera y más a un árbol irregular, con ramas que brotan, se cruzan, se extinguen o continúan.
También el medio ambiente desempeñó un papel decisivo en este proceso. Los cambios climáticos y la inestabilidad ecológica del Plio-Pleistoceno favorecieron nuevos paisajes y exigieron respuestas adaptativas variadas. En ese contexto, la flexibilidad alimentaria, la movilidad, el uso creciente de herramientas y la capacidad de ocupar entornos diversos debieron de influir en la diferenciación de las primeras especies humanas. La evolución del género Homo no puede entenderse solo como una serie de cambios anatómicos, sino como una respuesta compleja a condiciones ambientales cambiantes.
En conjunto, el recorrido por las primeras formas de Homo muestra hasta qué punto resulta difícil fijar un único origen del “aspecto humano”. Más que un momento preciso, lo que encontramos es una fase de transición prolongada, con especies cercanas entre sí, rasgos compartidos y caminos evolutivos parcialmente divergentes. Esa complejidad, lejos de ser un problema secundario, es precisamente una de las claves para entender nuestra historia: el ser humano no surgió como una figura acabada, sino como el resultado provisional de una evolución rica, abierta y profundamente dinámica.
Referencias
- Smithsonian Institution, Human Origins Program
- Encyclopaedia Britannica, Homo / Human evolution
- Baab et al., New reconstruction of DAN5 cranium (Gona, Ethiopia), Nature Communications (2025)
- Timmermann et al., Climate effects on archaic human habitats and species, Nature (2022)

Evolución biológica del género Homo
Cualquier reconstrucción de la evolución humana es necesariamente provisional. Las teorías cambian a medida que aparecen nuevos fósiles, se revisan antiguas atribuciones o mejoran los métodos de datación y análisis genético. Por eso, más que una historia cerrada, la evolución del género Homo debe entenderse como una interpretación en marcha, sometida a revisión constante. Aun así, sí puede trazarse una visión general razonablemente aceptada por buena parte de la comunidad científica.
Durante mucho tiempo se pensó que las industrias olduvayenses más antiguas estaban representadas por los útiles hallados en Gona, en Etiopía, datados entre 2,6 y 2,5 millones de años. Esos hallazgos siguen siendo fundamentales para entender el inicio de la tecnología lítica. Sin embargo, investigaciones más recientes han propuesto conjuntos aún más antiguos en África oriental, de modo que hoy resulta más prudente afirmar que Gona representa uno de los primeros grandes testimonios bien datados de la talla de piedra, pero no necesariamente el único ni el más antiguo en sentido absoluto.
También en Etiopía, en Hadar, apareció el maxilar A.L. 666-1, descrito a partir de un hallazgo realizado en 1994 y datado en torno a 2,33 millones de años. Sus rasgos han llevado a varios especialistas a relacionarlo con formas tempranas del género Homo, lo que lo convierte en una pieza muy importante para estudiar la transición entre los australopitecos tardíos y los primeros humanos. Aun así, como ocurre con otros fósiles muy antiguos, su interpretación no ha estado exenta de debate.
Una de las hipótesis que se han propuesto para explicar el aumento del cerebro en los primeros representantes del género Homo es la llamada teoría del tejido costoso, formulada por Leslie Aiello y Peter Wheeler. Según esta idea, el crecimiento del encéfalo, muy exigente desde el punto de vista energético, pudo ir acompañado de una reducción relativa del aparato digestivo, favorecida por una dieta de mayor calidad y más fácil de procesar. En este marco, el mayor consumo de carne, médula y otros alimentos energéticos habría contribuido a sostener un cerebro más grande. No obstante, esta hipótesis se considera hoy una propuesta influyente, pero no una explicación única ni definitiva del proceso.
En términos cronológicos, Homo habilis ha sido considerado durante mucho tiempo uno de los primeros miembros claros del género Homo. Su nombre, “hombre hábil”, refleja la asociación tradicional con la fabricación o el uso sistemático de útiles de piedra. Sin embargo, hoy se sabe que el panorama fue más complejo. Las primeras tecnologías líticas pudieron aparecer antes o al margen de Homo habilis, y la frontera entre este y otros homininos contemporáneos no siempre resulta nítida. Aun así, Homo habilis sigue ocupando un lugar importante como una de las formas humanas más antiguas ampliamente reconocidas, con un cerebro algo mayor que el de los australopitecos y una anatomía todavía bastante arcaica.
La idea clásica según la cual Homo habilis habría evolucionado de forma lineal hacia Homo erectus ha sido revisada con el tiempo. Hoy se tiende a pensar que entre hace algo más de dos millones y alrededor de 1,8 millones de años coexistieron varias especies tempranas del género Homo, entre ellas Homo habilis, Homo rudolfensis y las primeras formas relacionadas con Homo erectus o Homo ergaster. Esta coexistencia rompe la imagen simple de una sucesión ordenada y refuerza la idea de una evolución en mosaico, con varias ramas solapadas en el tiempo.
Homo erectus fue, en cualquier caso, una de las especies más exitosas de toda la evolución humana. Surgió en África y acabó extendiéndose por amplias regiones del Viejo Mundo, desde el Cáucaso y Asia occidental hasta China e Indonesia. Durante mucho tiempo se le consideró el gran protagonista de la primera expansión humana fuera de África. Más tarde, distintas poblaciones arcaicas derivadas de ese fondo evolutivo fueron dando lugar a formas regionales más recientes, algunas de las cuales suelen agruparse bajo denominaciones como Homo heidelbergensis o formas arcaicas de Homo sapiens.
Los hallazgos de la Sima de los Huesos, en Atapuerca, junto con los estudios genéticos de las últimas décadas, han reforzado la idea de que la evolución humana reciente no siguió una sola línea. De forma muy resumida, suele hablarse de una rama occidental, desarrollada en Europa y parte de Asia, que condujo a los neandertales, y de otra africana, relacionada con el surgimiento de Homo sapiens. Esta visión general sigue abierta a discusión en sus detalles, pero ha permitido abandonar los viejos esquemas demasiado rígidos y entender mejor la diversidad de poblaciones humanas del Pleistoceno.
En ese contexto, la hipótesis del origen africano reciente de Homo sapiens cuenta hoy con un fuerte respaldo genético y fósil. Según esta interpretación, nuestra especie surgió en África y desde allí se expandió al resto del mundo. Esta visión se opone al antiguo modelo multirregional, que proponía una aparición paralela de Homo sapiens en distintas partes del Viejo Mundo. No obstante, el panorama actual incorpora un matiz importante: la expansión de Homo sapiens no supuso una sustitución completamente aislada, ya que hubo cruces con otras humanidades, como neandertales y denisovanos.
El caso de los denisovanos ha sido especialmente revelador. A partir del estudio del ADN antiguo se ha podido identificar una población humana distinta de los neandertales y de los sapiens, presente en Asia durante el Pleistoceno tardío. En los últimos años, nuevos análisis genéticos y proteómicos han permitido relacionar con los denisovanos algunos fósiles asiáticos antes difíciles de clasificar, lo que está cambiando de forma notable nuestra visión de la evolución humana en el este de Eurasia.
Algo parecido ocurre con Homo floresiensis. Cuando se descubrió, muchos investigadores pensaron que podía tratarse de una forma insular derivada de Homo erectus. Más tarde surgieron hipótesis alternativas, según las cuales su origen podría remontarse a un linaje humano más antiguo. Hoy sigue sin haber un consenso absoluto sobre su posición exacta, pero su existencia demuestra hasta qué punto la evolución del género Homo fue diversa, irregular y llena de experimentos adaptativos. También otros fósiles tardíos de Asia, difíciles de encajar en categorías clásicas, recuerdan que el mapa humano reciente fue mucho más complejo de lo que se creyó durante décadas.
En conjunto, la evolución biológica del género Homo no puede describirse ya como una escalera recta que conduce inevitablemente hasta nosotros. Fue un proceso ramificado, con especies coexistentes, migraciones sucesivas, mezclas genéticas, adaptaciones locales y extinciones. Precisamente por eso resulta tan difícil fijar una frontera exacta entre los primeros Homo y otros homininos anteriores. Más que una aparición repentina, lo que vemos es una larga zona de transición en la que fueron emergiendo, poco a poco, algunos de los rasgos que hoy identificamos como humanos.
Referencias principales
- Semaw, S. et al., 2.5-million-year-old stone tools from Gona, Ethiopia, Nature (1997).
- Kimbel, W. H., Johanson, D. C. y Rak, Y., Systematic assessment of a maxilla of Homo from Hadar, Ethiopia, Journal of Human Evolution (1997).
- Aiello, L. C. y Wheeler, P., The Expensive-Tissue Hypothesis: The Brain and the Digestive System in Human and Primate Evolution, Current Anthropology (1995).
- Smithsonian Institution, Human Origins Program.
- Marshall, M., Who were the ancient Denisovans?, Nature (2025).
Especies del género Homo más aceptadas
- Homo habilis — ca. 2,4–1,65 millones de años
Una de las primeras especies del género Homo. Conserva bastantes rasgos arcaicos, pero ya muestra un cerebro algo mayor que el de los australopitecos y suele asociarse al uso temprano de herramientas de piedra. - Homo rudolfensis — ca. 1,9–1,8 millones de años
Especie africana temprana, conocida sobre todo por fósiles de la cuenca del Turkana. Suele distinguirse de H. habilis por su cara más ancha y algunos rasgos dentales y craneales, aunque la separación entre ambas sigue discutiéndose. - Homo ergaster — ca. 1,9–1,4 millones de años
Muchos investigadores lo consideran la forma africana temprana de Homo erectus; otros lo mantienen como especie separada. Representa un humano ya más alto, más proporcionado y con locomoción plenamente eficiente, muy importante en la expansión inicial fuera de África. - Homo erectus — ca. 1,89 millones–110.000 años
Es una de las especies humanas más exitosas y duraderas. Fue la primera en expandirse ampliamente por Eurasia y muestra una anatomía ya claramente humana en proporciones corporales, marcha y adaptación ecológica. - Homo antecessor — ca. 900.000–800.000 años
Conocido sobre todo por los fósiles de Atapuerca. Es una especie europea muy importante, aunque su posición exacta en el árbol evolutivo sigue siendo debatida; suele verse como una forma humana antigua de Europa occidental. - Homo heidelbergensis — ca. 700.000–200.000 años
Gran especie del Pleistoceno medio, con amplia distribución en África y Eurasia. Suele considerarse próxima al origen de neandertales en Europa y también relacionada con la línea que conduce a Homo sapiens en África. - Homo naledi — ca. 335.000–236.000 años
Descubierto en Sudáfrica, combina rasgos muy primitivos con otros sorprendentemente modernos. Su pequeño cerebro y su datación relativamente reciente lo convierten en una de las especies más desconcertantes del género Homo. - Homo neanderthalensis — ca. 400.000–40.000 años
Humanidad propia de Europa y parte de Asia occidental. Tenía gran capacidad craneal, una fuerte adaptación al clima frío y coexistió con Homo sapiens, con quien además se cruzó biológicamente. - Homo sapiens — desde ca. 300.000 años hasta hoy
Nuestra especie. Surgió en África y después se expandió por el resto del mundo. Es la única especie humana superviviente en la actualidad. - Homo floresiensis — ca. 100.000–50.000 años
La célebre especie de Flores, en Indonesia, de pequeña talla y cerebro reducido. Su posición exacta sigue discutiéndose, pero su validez como especie es ampliamente aceptada. - Homo luzonensis — ca. 67.000–50.000 años
Especie insular descubierta en Luzón, Filipinas. Presenta una combinación singular de rasgos dentales y óseos, y muestra que la diversidad humana tardía en Asia fue mayor de lo que antes se creía. - Nombres útiles, pero hoy más discutidos o no universalmente aceptados
- Homo georgicus — ca. 1,8 millones de años
Nombre usado para los fósiles de Dmanisi, en Georgia. Muchos especialistas prefieren incluirlos dentro de Homo erectus temprano, así que conviene tratarlo como un nombre útil pero no imprescindible en un listado depurado. - Homo cepranensis — ca. 430.000–350.000 años
Propuesto para el cráneo de Ceprano, en Italia. Hoy suele interpretarse más bien como una forma arcaica de Homo heidelbergensis que como una especie bien separada. - Homo rhodesiensis — ca. 324.000–274.000 años
Nombre clásico asociado al cráneo de Kabwe, en Zambia. Sigue usándose en algunos contextos, pero muchos autores prefieren incluir ese material dentro de Homo heidelbergensis o de formas africanas arcaicas cercanas a Homo sapiens. - Homo helmei — ca. 259.000 años
Nombre histórico vinculado al cráneo de Florisbad, en Sudáfrica. Hoy suele verse más como una denominación antigua para fósiles que se interpretan mejor como Homo heidelbergensis tardío o Homo sapiens temprano. - Denisovanos — ca. 200.000–40.000 años
Grupo humano bien real y muy importante, conocido sobre todo por genética y algunos fósiles de Asia. El problema es que “Homínido de Denísova” no tiene todavía una denominación binomial universalmente aceptada como especie formal del tipo Homo xxxxx. - Hombres de la cueva de los ciervos — ca. 14.000–11.000 años
Población humana del suroeste de China muy discutida. No se acepta de forma general como especie separada del género Homo; suele tratarse como un conjunto fósil problemático y todavía abierto a interpretación.
Nombres que yo quitaría de un listado principal, salvo nota aparte
- Homo gautengensis
Es una propuesta de 2010, pero ha tenido muy poca aceptación. La mayor parte del material asignado a este nombre sigue repartiéndose entre H. habilis, H. erectus, H. ergaster u otros taxones. - Homo tsaichangensis
No es un nombre asentado en el consenso general ni aparece entre las especies aceptadas por los grandes resúmenes de referencia. Yo no lo pondría en un listado principal de tu blog, salvo quizá en una nota sobre propuestas muy recientes o muy controvertidas.
- Smithsonian Institution, Human Origins Program
- Nature, A new species of Homo from the Late Pleistocene of the Philippines (2019)
- Nature, Who were the ancient Denisovans? Fossils reveal secrets about the mysterious humans (2025)
- Scientific Reports, Digital reconstruction of the Ceprano calvarium (2017)
- Science, A male Denisovan mandible from Pleistocene Taiwan (2025)
No todas las especies atribuidas al género Homo gozan del mismo grado de aceptación. En paleoantropología, algunas denominaciones están muy consolidadas, mientras que otras siguen siendo provisionales, discutidas o dependen de fósiles demasiado escasos como para establecer conclusiones firmes. Por eso conviene distinguir entre las especies generalmente admitidas y los nombres que se utilizan con más cautela, ya sea por falta de restos, por problemas de clasificación o porque podrían representar variantes regionales de especies ya conocidas.
Uno de los casos clásicos es el de Homo ergaster. Para muchos investigadores, se trata de una especie africana bien diferenciada y anterior a Homo erectus; para otros, en cambio, no sería más que la forma africana temprana de Homo erectus. Algo parecido ocurre con Homo georgicus, nombre propuesto para los célebres fósiles de Dmanisi, en Georgia: algunos especialistas lo aceptan como especie propia, mientras que otros prefieren incluir esos restos dentro de un Homo erectus muy primitivo. En ambos casos, el debate gira en torno a una cuestión de fondo: si conviene multiplicar las especies o si es mejor interpretar esas diferencias como parte de la variabilidad de un linaje humano amplio y antiguo.
Más problemáticos todavía son algunos nombres basados en restos muy fragmentarios. Homo cepranensis, propuesto a partir del cráneo de Ceprano, en Italia, no ha logrado un consenso pleno y con frecuencia se interpreta como una forma cercana a Homo heidelbergensis. Algo semejante sucede con Homo rhodesiensis, nombre histórico asociado sobre todo al cráneo de Kabwe, en Zambia: muchos autores lo siguen utilizando, pero otros prefieren verlo como una variante africana de Homo heidelbergensis o como una forma arcaica próxima al origen de Homo sapiens. Homo helmei, por su parte, ha tenido un uso aún más restringido y suele entenderse más como una propuesta hipotética para ciertos fósiles africanos de transición que como una especie firmemente establecida.
En otros casos, las discusiones afectan a especies insulares o tardías. Homo floresiensis, descubierto en la isla de Flores, generó en su momento una gran controversia, y durante años algunos investigadores defendieron que podía tratarse de un Homo sapiens patológico. Hoy, sin embargo, la posición mayoritaria lo reconoce como un taxón válido y distinto de nuestra especie, aunque siga abierto el debate sobre su origen exacto: si desciende de Homo erectus, de una forma humana más primitiva o de una dispersión muy antigua aún mal documentada.
El caso de los neandertales también merece una precisión. Durante mucho tiempo fueron tratados a veces como una subespecie, Homo sapiens neanderthalensis, pero hoy la tendencia dominante los considera una especie distinta, Homo neanderthalensis. Los estudios genéticos han reforzado esa distinción, aunque también han mostrado que neandertales y Homo sapiens se cruzaron entre sí. Por tanto, no eran simples variantes de la humanidad actual, sino una humanidad paralela, cercana, compatible biológicamente en parte, pero diferenciada.
A esto se suma el problema de los denisovanos. Se trata de un grupo humano bien real, conocido gracias al ADN antiguo y a algunos fósiles de Asia, pero que todavía no posee una denominación binomial universalmente asentada dentro del género Homo. Durante años fueron casi una “humanidad fantasma”, identificable sobre todo por genética; hoy su realidad biológica está fuera de duda, pero su encaje taxonómico exacto sigue abierto. Algo parecido, aunque con mucha menor solidez, ocurre con los llamados hombres de la cueva de los ciervos, en China, cuya posición permanece incierta y no justifica, por ahora, tratarlos como una especie bien establecida.
En los últimos años se añadió además el nombre Homo longi, propuesto en 2021 a partir del cráneo de Harbin, en China. Su presentación despertó mucho interés porque algunos autores lo consideraron una nueva especie importante dentro de la evolución humana asiática. Sin embargo, trabajos posteriores han discutido esa interpretación, y estudios proteómicos recientes han sugerido que el individuo de Harbin podría pertenecer en realidad al linaje denisovano. Por eso, hoy lo más prudente es tratar Homo longi como una propuesta discutida, todavía lejos de contar con una aceptación general.
Algo similar vale para el nombre Homo tsaichangensis, que se ha relacionado con una mandíbula fósil hallada en Taiwán. El problema es que los estudios más influyentes publicados recientemente no refuerzan la necesidad de crear una especie nueva, sino que apuntan más bien a una afinidad denisovana para ese fósil. En consecuencia, incluir Homo tsaichangensis como especie consolidada resultaría prematuro.
En conjunto, todo esto muestra que la clasificación del género Homo está lejos de ser completamente estable. Algunos nombres perduran porque resultan útiles para ordenar materiales fósiles diversos, mientras que otros podrían acabar desapareciendo o integrándose en especies ya conocidas. La paleoantropología trabaja con restos escasos, cronologías amplias y poblaciones que a menudo compartieron rasgos, territorios e incluso herencia genética. Por eso, más que una lista cerrada y definitiva, el género Homo debe entenderse como un campo en revisión continua, en el que cada nuevo hallazgo puede obligar a reorganizar el mapa.
Referencias principales
- Smithsonian Institution, Human Origins Program
- Michael Marshall, “Who were the ancient Denisovans? Fossils reveal secrets about the mysterious humans”, Nature (2025)
- Fu et al., estudio proteómico sobre el cráneo de Harbin y su relación con los denisovanos, Science (2025)
- Tsutaya et al., estudio sobre la mandíbula pleistocena de Taiwán y su afinidad denisovana, Science (2025)
- Estudios de síntesis recientes sobre Homo floresiensis y la diversidad humana de Asia oriental
Referencias
- Wood, B. (1992): «Origin and evolution of the genus Homo». Nature, 355: 783-790
- Mann, A. y Weiss, M. (1996): «Hominoid Phylogeny and Taxonomy : a consideration of the molecular and Fossil Evidence in an Historical Perspective». Molecular Phylogenetics and Evolution, 5(1): 169-181
- Sutikna, Thomas; Tocheri, Matthew W.; Morwood, Michael J.; Saptomo, E. Wahyu; Jatmiko; Awe, Rokus Due; Wasisto, Sri; Westaway, Kira E. et al. (21 de abril de 2016). «Revised stratigraphy and chronology for Homo floresiensis at Liang Bua in Indonesia». Nature (en inglés) 532 (7599): 366-369.
- Leakey, R.E. (1981): «The Making of Mankind». New York: E.P. Dutton, p.p. 65-66. ISBN 0-525-15055-2
- SÁEZ, Roberto (2019). Evolución Humana: Prehistoria y origen de la compasión (en espanyol). Espanya: Almuzara. p. 51.
- Arsuaga, J.L. y equipo: Claves de la evolución humana
- Semaw, S.; P. Renne; W. K. J Harris; C. S. Feibel; R. L. Bernor; N. Fesseha & K. Mowbray (1997) «2.5-million-year-old stone tools from Gona, Ethiopia»; Nature 385 (6614): 333-336.
- Kimbel, William H.; Donald C. Johanson & Yoel Rak (1998) «Systematic assessment of a maxilla of Homo from Hadar, Ethiopia»; American Journal of Physical Anthropology 103 (2): 235–262.
- Ciudad Universitaria Virtual de San Isidoro. «La teoría del tejido costoso». Consultado el 22 de octubre de 2016.
- Leakey, L.S.B.; Tobias, P.V. y Napier, J.R. (1964) «A New Species of Genus Homo from Olduvai Gorge» (en inglés»; Nature 202 (4927): 7-9.
- Rightmire, G.P. (1990): The Evolution of Homo erectus. Comparative Anatomical Studies of an Extinct Human Species. Cambridge: Cambridge University Press.
- Tres especies de Homo distintas convivieron hace dos millones de años Tendencias científicas, agosto de 2012
- Serre et al. (2004): «No evidence of Neandertal mtDNA contribution to early modern humans». PLoS Biology, 2(3): 313–7
- Rightmire, G.P. (1992): «Homo erectus: ancestor or evolutionary side branch?». Evolutionary Anthropology, 1: 43-4
- Maca-Meyer, N., González, A.M., Larruga, J.M., Flores, C. y Cabrera, V.M. (2001): «Linajes mayores del genoma mitocondrial trazan antiguas expansiones humanas». BMC Genetics, 2: 13
- Ruiz, H. (2007): «El origen de los humanos modernos«. Eureka, 11: 36-41
- Krause, Johannes; Fu, Qiaomei; Good, Jeffrey M.; Viola, Bence; Shunkov, Michael V.; Derevianko, Anatoli P. & Pääbo, Svante (2010), «The complete mitochondrial DNA genome of an unknown hominin from southern Siberia»; Nature 464 (7290): 894–897.
- Reich, David; Richard E. Green, et.al. (22 December, 2010) «Genetic history of an archaic hominin group from Denisova Cave in Siberia»; Nature 468 (1012): 1053–1060.
- Brown, P. et al. (2004): «A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia». Nature, 431: 1055-1061
- Jungers, William L.; W. E. H. Harcourt-Smith; R. E. Wunderlich; M. W. Tocheri; S. G. Larson; T. Sutikna; Rhokus Awe Due & M. J. Morwood (2009) «The foot of Homo floresiensis»; Nature 459: 81-84.
- Homo ergaster Archivado el 12 de marzo de 2008 en Wayback Machine.; Archaelogiy.info.
- Spoor, F.; Leakey, M. G.; Gathogo, P. N.; Brown, F. H.; Antón, S. C.; McDougal, I.; Kiarie, C.; Manthi, F. K. y Leakey, L. N. (2007) «Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya». Nature, 448: 688-691.
- Rightmire, G. Philip (1998). «Human evolution in the Middle Pleistocene: The role of Homo heidelbergensis»; Evolutionary Anthropology 6: 218-227.
- G. Philip Rightmire (2009) Middle and later Pleistocene hominins in Africa and Southwest Asia; PNAS.
- Hershkovitz, Israel; Konreich, L; Laron, Z (2007). «Comparative Skeletal Features Between Homo floresiensis and Patients With Primary Growth Hormone Insensitivity (Laron Syndrome)». American Journal of Physical Anthropology (en inglés). Archivado desde el original el 11 de enero de 2012.
En este video respondemos a algunas de las objeciones que nos han dejado en la sección de comentarios del video «Teoría de la evolución», y hablamos de la aparición de nuevas especies y de la hominización, que es el proceso evolutivo que llevó al desarrollo del ser humano y sus parientes cercanos extintos. Agradecemos a quienes puedan colaborar con el mantenimiento de este canal a través de Patreon, Paypal (opción pago por servicio o producto, NO opción pago personal) y Mercado pago. Patreon
 / roxanakreimer
/ roxanakreimer Epílogo: balance final sobre el género Homo
Al llegar al final de un recorrido por el género Homo, lo primero que se impone no es una sensación de simplicidad, sino todo lo contrario: la conciencia de haber contemplado una historia enormemente rica, cambiante y llena de matices. Durante mucho tiempo se presentó la evolución humana como una secuencia lineal, casi escolar, en la que una especie sucedía limpiamente a otra hasta desembocar en el ser humano actual. Hoy sabemos que aquella imagen era demasiado pobre. El género Homo no fue una escalera ascendente, sino un conjunto de ramas, tanteos, adaptaciones y coexistencias. Hubo especies que perduraron durante cientos de miles de años, otras que apenas conocemos por unos pocos restos, algunas que se expandieron por continentes enteros y otras que quedaron aisladas en espacios concretos. El resultado general no fue una marcha uniforme hacia nosotros, sino una auténtica pluralidad de humanidades.
Esa diversidad obliga a mirar el género Homo con una mezcla de admiración y prudencia. Admiración, porque en él se concentran algunos de los cambios más decisivos de toda la evolución de los homininos: el aumento del cerebro, el perfeccionamiento de la marcha bípeda, la creciente complejidad técnica, la ampliación de la dieta, la expansión geográfica y, con el tiempo, la aparición de conductas simbólicas y culturales de enorme alcance. Prudencia, porque ninguno de esos cambios surgió de golpe ni apareció de forma igual en todas las poblaciones. La evolución del género Homo fue, en gran medida, una evolución en mosaico: unos grupos desarrollaron antes determinadas capacidades, otros conservaron rasgos arcaicos, y muchos compartieron combinaciones anatómicas y conductuales difíciles de encajar en categorías rígidas. Por eso cada nuevo fósil, cada nueva datación y cada nueva técnica de análisis obligan a revisar una parte del mapa.
Uno de los aspectos más impresionantes de este proceso es su profundidad temporal. El género Homo se remonta, según la interpretación más aceptada, a una franja cercana a los 2,8–2,5 millones de años, aunque el registro de esos primeros momentos siga siendo incompleto y discutido. Desde aquellas formas tempranas hasta la aparición de Homo sapiens en África hace más de 300.000 años, se extiende una larga etapa de cambios graduales, bifurcaciones y ensayos evolutivos. En ese tiempo aparecen especies tempranas como Homo habilis y Homo rudolfensis; más tarde, formas de gran éxito adaptativo como Homo erectus; después, poblaciones del Pleistoceno medio relacionadas con Homo heidelbergensis y otros grupos afines; y finalmente, humanidades más próximas a nosotros, como neandertales, denisovanos y los primeros sapiens. Esa cronología no dibuja un relevo ordenado, sino un panorama donde varias especies pudieron convivir y hasta solaparse durante largos periodos.
También resulta decisivo comprender que el género Homo no puede explicarse solo por la anatomía. Su historia está ligada a cambios ecológicos profundos y a nuevas formas de relación con el medio. Las oscilaciones climáticas del Plio-Pleistoceno, la expansión de paisajes abiertos, la necesidad de recorrer mayores distancias, el acceso a nuevos recursos y la fabricación de herramientas influyeron poderosamente en la configuración de estas especies. A medida que avanzó el tiempo, los miembros del género Homo fueron mostrando una flexibilidad cada vez mayor para adaptarse a ambientes distintos. Algunos grupos prosperaron en las sabanas africanas; otros ocuparon regiones templadas o frías de Eurasia; otros lograron sobrevivir en islas del sudeste asiático. Esta capacidad de diversificación ecológica fue una de las claves de su éxito.
Entre todas las especies del género, Homo erectus ocupa un lugar especial. No solo por su antigüedad y duración, sino porque representa uno de los grandes saltos de la evolución humana. Con él aparecen ya unas proporciones corporales claramente modernas, una expansión geográfica sin precedentes y una notable estabilidad evolutiva a lo largo de muchísimo tiempo. Fue, en cierto modo, la primera humanidad verdaderamente cosmopolita del Viejo Mundo. Sin embargo, ni siquiera Homo erectus puede entenderse como un punto de llegada definitivo. Su legado se ramificó, dio lugar a nuevas poblaciones y dejó abierta la historia posterior de Europa, Asia y África. En ese sentido, más que una especie cerrada, fue una plataforma evolutiva de enorme alcance.
Otro gran aprendizaje que deja el estudio del género Homo es la caída de cualquier idea simple de excepcionalidad inmediata. Durante mucho tiempo tendimos a imaginarnos como el destino inevitable de toda la secuencia anterior, casi como si las demás especies hubieran existido para conducir finalmente hasta nosotros. Hoy esa visión resulta insuficiente. Los neandertales no fueron un borrador defectuoso del ser humano moderno, ni los denisovanos una nota al pie, ni Homo floresiensis o Homo luzonensis simples rarezas marginales. Todos ellos fueron humanidades reales, adaptadas a sus circunstancias, con historias propias y, en algunos casos, con logros biológicos y culturales notables. Nuestra especie no es el sentido oculto de toda la evolución anterior, sino una de las ramas que finalmente sobrevivió. Entender esto aporta una mirada más humilde y más verdadera sobre la condición humana.
La genética ha reforzado de manera espectacular esta nueva visión. Gracias al ADN antiguo sabemos que la historia del género Homo no fue solo una sucesión de reemplazos, sino también una historia de encuentros, mezclas y herencias compartidas. Homo sapiens se cruzó con neandertales y con denisovanos, y una parte de esas huellas sigue presente hoy en poblaciones humanas actuales. Esto ha derribado la vieja oposición entre especies absolutamente incomunicadas y ha mostrado que la frontera entre unas humanidades y otras fue, en algunos momentos, más porosa de lo que se suponía. Al mismo tiempo, los estudios recientes sobre los denisovanos y sobre fósiles asiáticos como el cráneo de Harbin están obligando a repensar la diversidad humana del este de Eurasia, una región que probablemente fue mucho más compleja de lo que dejaban ver los esquemas tradicionales.
En el fondo, el estudio del género Homo tiene una fuerza especial porque nos sitúa ante una doble evidencia. Por un lado, nos muestra que pertenecemos plenamente al mundo natural: somos el resultado de una historia biológica larguísima, compartimos ancestros con otras especies y llevamos en el cuerpo las huellas de esa procedencia evolutiva. Por otro lado, también nos recuerda que dentro de esa historia natural surgieron capacidades singulares: técnica acumulativa, lenguaje articulado, memoria simbólica, transmisión cultural compleja, conciencia histórica. No aparecieron de repente ni como un milagro aislado, sino como culminación provisional de una larga trayectoria de cambios graduales. Lo humano, visto desde esta perspectiva, no rompe con la naturaleza, sino que brota de ella y la transforma.
Por eso un artículo sobre el género Homo no termina de verdad con una simple enumeración de especies, fechas o yacimientos. Termina, más bien, con una pregunta de fondo: qué significa ser humano cuando sabemos que no hemos estado solos, que hemos compartido el mundo con otras humanidades y que nuestra propia forma actual es solo el último capítulo conocido de una historia mucho más amplia. Estudiar el género Homo no sirve únicamente para reconstruir un pasado remoto; sirve también para descentrarnos, para relativizar nuestra aparente autosuficiencia y para comprender que llevamos dentro una memoria biológica mucho más antigua que cualquier civilización. En ese sentido, la paleoantropología no solo ilumina nuestros orígenes: también ensancha nuestra idea de nosotros mismos.
Y quizá ahí resida la grandeza última de este tema. El género Homo no es solo una clasificación zoológica ni una serie de nombres latinos. Es el relato de una aventura biológica extraordinaria, llena de avances, pérdidas, errores, mezclas y supervivencias. Es la historia de cuerpos que cambiaron, de cerebros que crecieron, de manos que tallaron piedra, de grupos que cruzaron paisajes inmensos y de especies que aparecieron y desaparecieron sin saber que algún día serían reconstruidas por otra de sus parientes. Nosotros somos esa especie superviviente, sí, pero no la única humanidad que existió, ni probablemente la medida absoluta de lo humano. Cerrar un trabajo sobre el género Homo es, por tanto, cerrar provisionalmente una reflexión sobre la pluralidad de nuestra propia estirpe. Y esa reflexión, lejos de empequeñecernos, nos vuelve más conscientes, más humildes y también más interesantes.
Ver artículo actualizado: «El origen del género Homo y la hominización.»
Referencias principales
- Smithsonian Institution, Human Origins Program.
- Encyclopaedia Britannica, Homo; Human evolution; Homo sapiens.
- Michael Marshall, “Who were the ancient Denisovans? Fossils reveal secrets about the mysterious humans”, Nature (2025).
- Qiaomei Fu et al., “The proteome of the late Middle Pleistocene Harbin individual”, Science (2025).
- Liam Nic Eoin, “Face to face with a Denisovan”, Nature Ecology & Evolution (2026).